
This article briefly describes how lectins function as key mediators of biological processes by recognizing and interpreting the cellular “sugar code.” It outlines the role of lectin–carbohydrate interactions in regulating hormone circulation, immune cell trafficking, pathogen recognition, and intracellular protein targeting. The article also highlights the involvement of lectins in major diseases and their growing importance as therapeutic targets. Finally, it explains the molecular basis of lectin specificity and multivalency, emphasizing their central role in cellular communication.
Lectin-Mediated Recognition in Cellular Communication
Lectins are specialized carbohydrate-binding proteins that recognize specific sugar patterns on cell surfaces and translate these patterns into biological signals. By interpreting this sugar code, lectins help control processes such as cell adhesion, immune responses, pathogen recognition, inflammation, and development. Understanding how lectins read and interpret the sugar code has become a major focus in modern glycobiology, with implications ranging from disease diagnosis to drug development. This article examines the role of lectins as molecular interpreters of cellular communication and their vital importance for life.
Lectin-Mediated Regulation of Circulating Glycoproteins
The carbohydrate components attached to circulating glycoproteins play a crucial role in determining how long these molecules remain in the bloodstream. Several peptide hormones contain N-linked oligosaccharides, whose terminal sugar structures serve as signals that regulate their removal from the circulation.
Luteinizing hormone and thyrotropin, two pituitary-derived hormones, possess N-linked oligosaccharides that terminate in the disaccharide GalNAc4S(β1→4)GlcNAc. This specific carbohydrate motif is recognized by lectin receptors present on hepatocytes in the liver. The binding of these hormones to hepatic lectins triggers their uptake and degradation, thereby lowering their concentration in the blood. As a result, the levels of these hormones show characteristic cyclic fluctuations.
Terminal sialic acid residues (Neu5Ac) present on many plasma glycoproteins serve a protective function by preventing their rapid clearance. Ceruloplasmin, a copper-binding serum glycoprotein, contains several oligosaccharide chains capped with Neu5Ac residues that shield it from hepatic uptake. Removal of these terminal sialic acids, possibly through the action of neuraminidases released by microorganisms or extracellular enzymes, exposes underlying galactose residues. Hepatocyte lectins specifically recognize these exposed galactose-containing glycans, leading to receptor-mediated endocytosis and degradation of the glycoprotein. This mechanism highlights how lectin–glycan recognition controls the lifespan of circulating hormones and serum glycoproteins.
Lectin Recognition in Erythrocyte Clearance
A comparable lectin-dependent process is believed to contribute to the removal of aged red blood cells from the circulation in mammals. Newly formed erythrocytes display membrane glycoproteins whose oligosaccharide chains terminate in sialic acid (Neu5Ac), which helps maintain their persistence in the bloodstream. Experimental studies have shown that when these terminal sialic acid residues are enzymatically removed in vitro using neuraminidase, and the modified cells are reintroduced into circulation, the desialylated erythrocytes are rapidly cleared within hours. In contrast, erythrocytes retaining their intact sialylated glycans remain in circulation for several days. This observation supports the idea that loss of terminal sialic acid exposes underlying sugar residues that are recognized by hepatic lectins, marking the cells for rapid removal.
Selectins and Lectin-Mediated Immune Cell Trafficking
Cell-surface lectins play a major role in the development and progression of several diseases, particularly through their involvement in immune cell movement and adhesion. A key group of membrane lectins, known as selectins, mediates cell–cell recognition during many physiological and pathological processes. One of their most important functions is guiding immune cells from the bloodstream into tissues at sites of infection or inflammation.
At inflamed or infected regions, endothelial cells lining capillaries express P-selectin on their surface. This lectin recognizes specific carbohydrate structures present on glycoproteins of circulating leukocytes. The interaction causes leukocytes to slow down and roll along the endothelial surface. Subsequently, integrin proteins on leukocytes bind to adhesion molecules on endothelial cells, firmly arresting the leukocytes and enabling their passage through the capillary wall into the affected tissue, where they initiate immune responses (Figure 1).
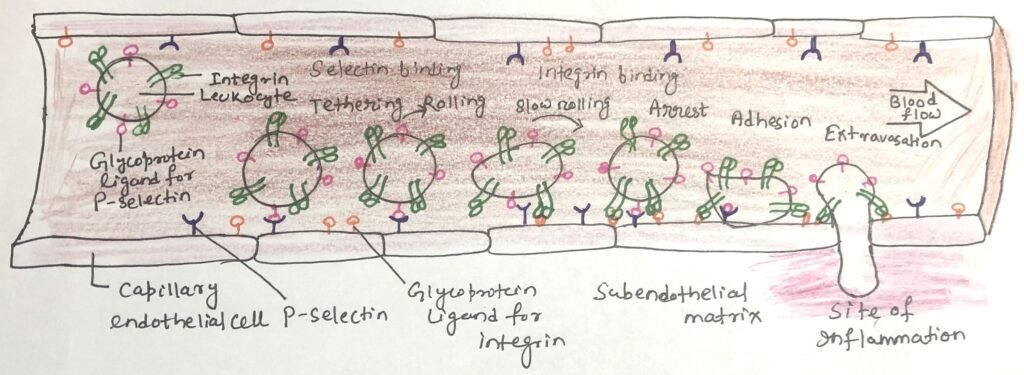
Additional members of the selectin family contribute to this process of lymphocyte homing. E-selectin, expressed by endothelial cells, and L-selectin found on leukocytes bind to their corresponding carbohydrate ligands on opposing cell surfaces, ensuring precise and regulated migration of immune cells.
Selectin-Driven Inflammatory Disorders and Tumor Metastasis
Selectin-mediated adhesion is also closely linked to the development and progression of several inflammatory and immune-related disorders. Excessive or misregulated selectin activity contributes to conditions such as rheumatoid arthritis, asthma, psoriasis, multiple sclerosis, and transplant rejection. Because of this, selectins have become important therapeutic targets, and considerable effort is being directed toward designing drugs that can block selectin-dependent cell adhesion and thereby reduce inflammation.
In addition, many carcinomas produce and release carbohydrate antigens that are typically found only during fetal development. When these antigens enter the bloodstream, they enhance the ability of tumor cells to evade immune defenses and promote metastatic spread. This highlights the broader pathological significance of lectin–glycan interactions in both inflammatory disease and cancer progression.
Lectin–Glycan Interactions in Viral Infection
Many animal viruses rely on carbohydrate recognition to initiate infection. Influenza virus, for example, attaches to host cells by binding to specific oligosaccharides present on the cell surface. This attachment is mediated by a viral lectin called hemagglutinin (HA), which plays a crucial role in viral entry.
Following replication inside the host cell, newly formed viral particles exit by budding from the plasma membrane, acquiring a portion of the host membrane as their envelope. During this stage, the viral enzyme neuraminidase (a sialidase) removes terminal sialic acid residues from host-cell glycans. This cleavage prevents newly formed viruses from remaining attached to the infected cell and reduces aggregation among viral particles, thereby enabling the viruses to spread efficiently and initiate new rounds of infection.
Lectins in Microbial Pathogenesis and Intracellular Protein Targeting
Many pathogenic microorganisms use lectin-mediated recognition to attach to host tissues or deliver toxins into cells. Helicobacter pylori, for instance, expresses a surface lectin that binds to specific oligosaccharides on the epithelial lining of the stomach. One of the key carbohydrate targets is the Lewis b (Leb) antigen. It is a structure present on glycoproteins and glycolipids associated with blood group O. This interaction helps explain why individuals with blood type O show a higher susceptibility to gastric ulcers. The bacterium can adhere more efficiently to the stomach epithelium. Synthetic molecules designed to mimic the Leb structure are being explored as therapeutic agents. This, when taken orally, could competitively block bacterial attachment by occupying the lectin-binding sites.
Several major parasitic diseases are also linked to organisms that display distinctive surface glycans. These surface glycans aid their survival and immune evasion. These include trypanosomes, which cause African sleeping sickness and Chagas disease. The other parasites are Plasmodium falciparum, responsible for malaria, and Entamoeba histolytica, which causes amoebic dysentery. The unusual carbohydrate coatings of these parasites often provide protection and enhance their ability to infect hosts. Lectins are not limited to extracellular interactions; they also function within cells by directing protein trafficking. A notable example is the mannose-6-phosphate tag found on certain newly synthesized proteins in the Golgi apparatus. Lectins recognize this carbohydrate marker and guide the tagged proteins to lysosomes, ensuring proper delivery to their functional destination.
Molecular Basis of the Sugar Code and Lectin Specificity
The structural diversity of oligosaccharides creates a highly information-rich system often described as the sugar code. This is capable of generating an enormous variety of distinct molecular “signals” that can be recognized by proteins. Lectins possess precisely shaped carbohydrate-binding sites that allow them to interact selectively with their correct glycan partners. This results in remarkable specificity. Although the binding strength between a single carbohydrate-binding domain (CBD) and an oligosaccharide may be relatively modest, lectins frequently contain multiple CBDs. This multivalency enables simultaneous interactions with clusters of glycans on cell surfaces, greatly enhancing the overall binding strength. When multiple lectin receptors are present on a cell, the combined interactions can become highly cooperative. These support processes, such as cell adhesion and rolling along vascular surfaces.
Structural studies, particularly X-ray crystallography of the mannose-6-phosphate receptor, have provided detailed insight into the basis of lectin–glycan recognition. These analyses reveal how the receptor specifically binds mannose-6-phosphate and how a divalent metal ion contributes to the interaction. For instance, the amino acid residue His105 forms a hydrogen bond with a phosphate oxygen, helping stabilize the complex. When the tagged protein reaches the lysosome, the receptor releases its ligand. Protonation of His105 is thought to reduce binding affinity and enable this release.
Beyond highly specific contacts, broader physicochemical forces also contribute to lectin–carbohydrate binding. Many sugars display both polar and nonpolar regions. The polar portions form hydrogen bonds with lectin residues, while the less polar regions interact with hydrophobic amino acids. The combined effect of these multiple interactions produces strong and selective binding. This illustrates how lectin–glycan recognition functions as an essential mechanism of information transfer within and between cells.
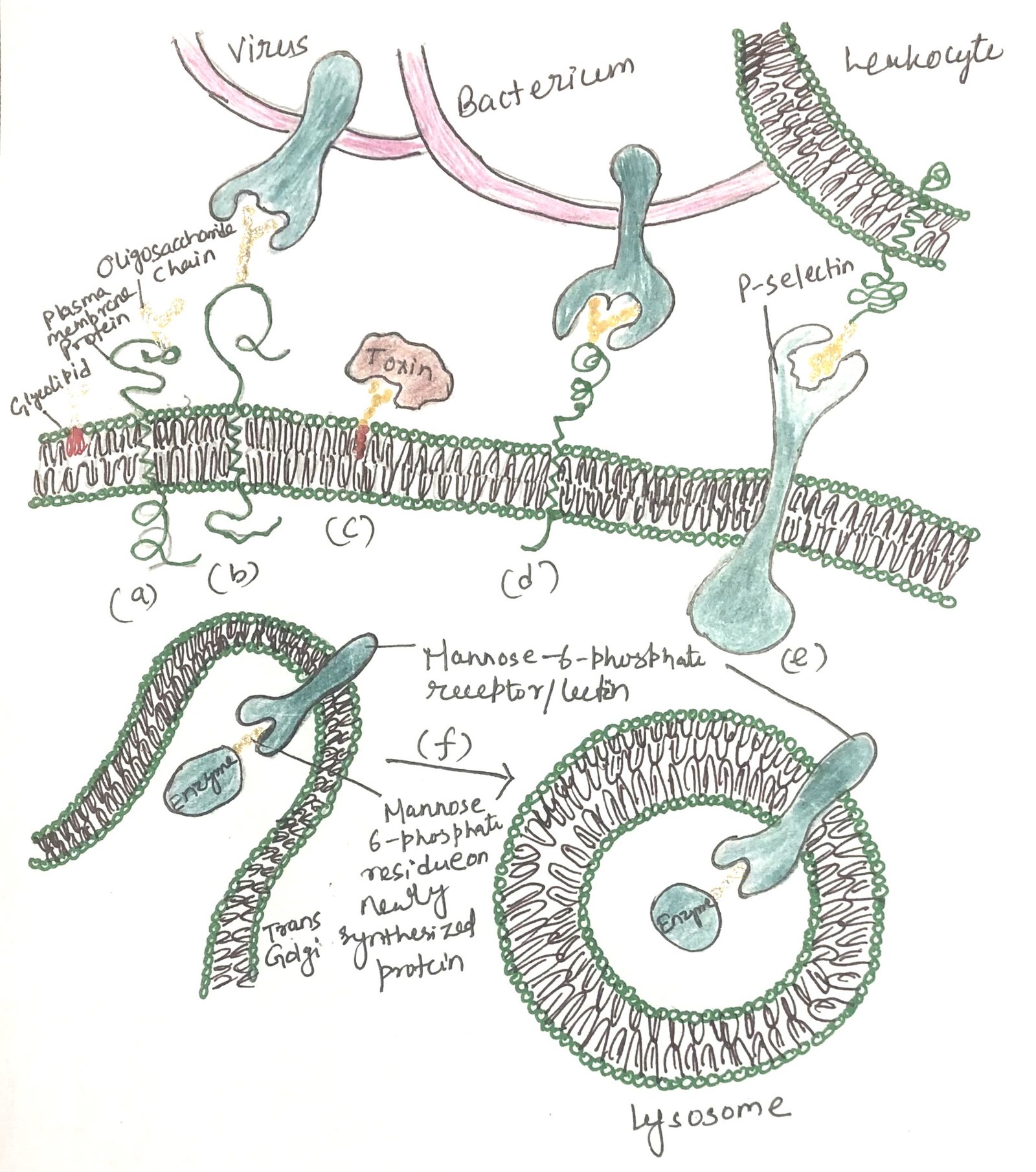
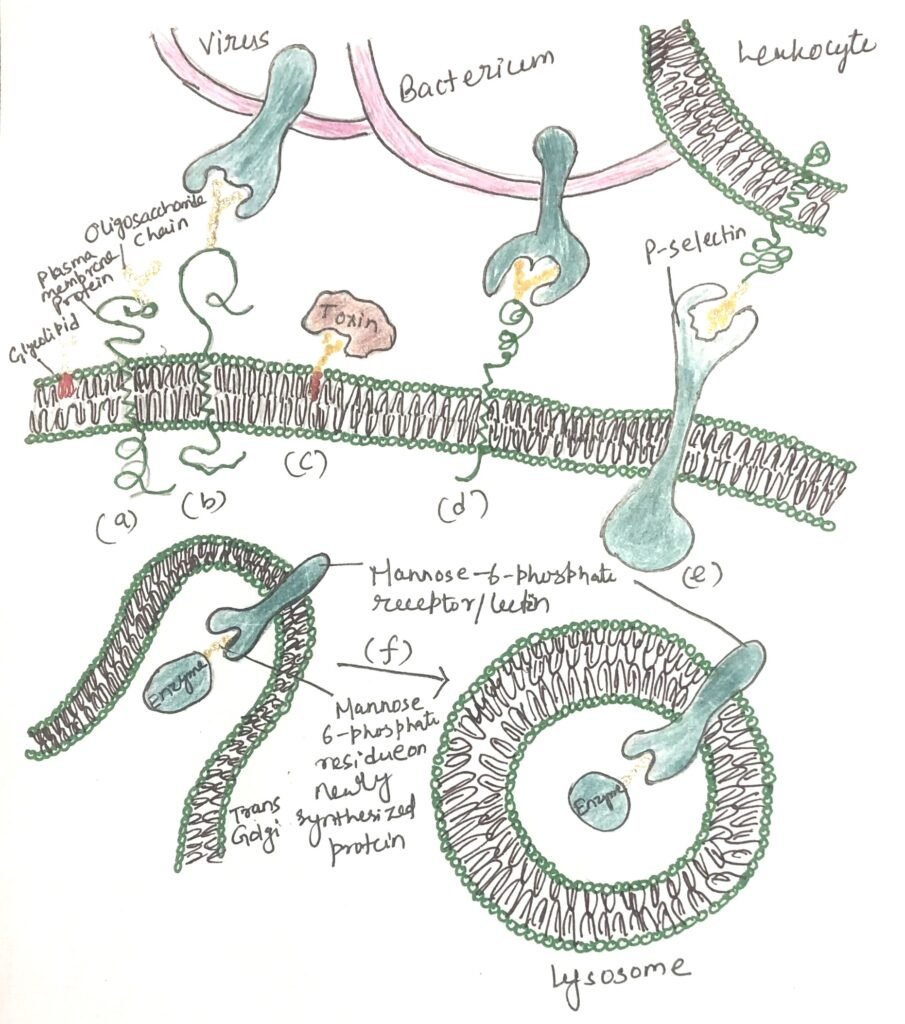
Sugar code mediates biological interactions
Distinctly structured oligosaccharides form part of many glycoproteins and glycolipids located on the external surface of the plasma membrane (Figure 2a). Their oligosaccharide moieties are bound by extracellular lectins with high specificity and affinity. Figure 2 shows a summary of the biological interactions mediated by the sugar code. Animal viruses, including the influenza virus, initiate infection by attaching to glycoproteins present on the host cell surface (Figure 2b). Certain bacterial toxins, including cholera and pertussis toxins, attach to surface glycolipids prior to gaining entry into cells (Figure 2c).
Some bacteria adhere to and then colonize or infect animal cells (Figure 2d). Selectins located in the plasma membrane of specific cells facilitate cell–cell interactions, including the binding of leukocytes to endothelial cells lining capillary walls at sites of infection (Figure 2e). The mannose-6-phosphate receptor in the trans-Golgi complex recognizes the oligosaccharides of lysosomal enzymes and directs them to the lysosome for delivery (Figure 2f).
Conclusion
Lectins serve as essential interpreters of the cellular sugar code, translating complex glycan patterns into biological responses that influence processes across nearly every level of life. From regulating the circulation of hormones and glycoproteins, guiding the clearance of aging erythrocytes, and directing immune cell trafficking, to enabling viral entry, microbial adhesion, and intracellular protein sorting, lectin–glycan interactions form a unifying mechanism of cellular communication. These interactions are characterized by remarkable specificity and enhanced binding through multivalency, allowing cells and pathogens alike to recognize precise carbohydrate signatures. Their involvement in inflammatory disorders, cancer progression, infectious diseases, and protein targeting highlights their profound medical relevance and therapeutic potential. As research in glycobiology continues to expand, understanding how lectins read and interpret the sugar code will remain central to developing new diagnostic tools and targeted treatments, underscoring the fundamental role of carbohydrate recognition in health and disease.
You may also like:

I, Swagatika Sahu (author of this website), have done my master’s in Biotechnology. I have around fourteen years of experience in writing and believe that writing is a great way to share knowledge. I hope the articles on the website will help users in enhancing their intellect in Biotechnology.