
This article briefly describes the types of cloning vehicles and their properties.
Cloning Vehicle
In a gene cloning experiment, a cloning vehicle plays a central role in transporting the gene into the host cell and is responsible for its replication. Plasmids and bacteriophages act as vectors for transporting genes. The vector carries the foreign DNA as the recombinant DNA. The factors that characterize the vectors can be defined as the origin of replication, multi-cloning site, and a selectable marker.
Plasmids
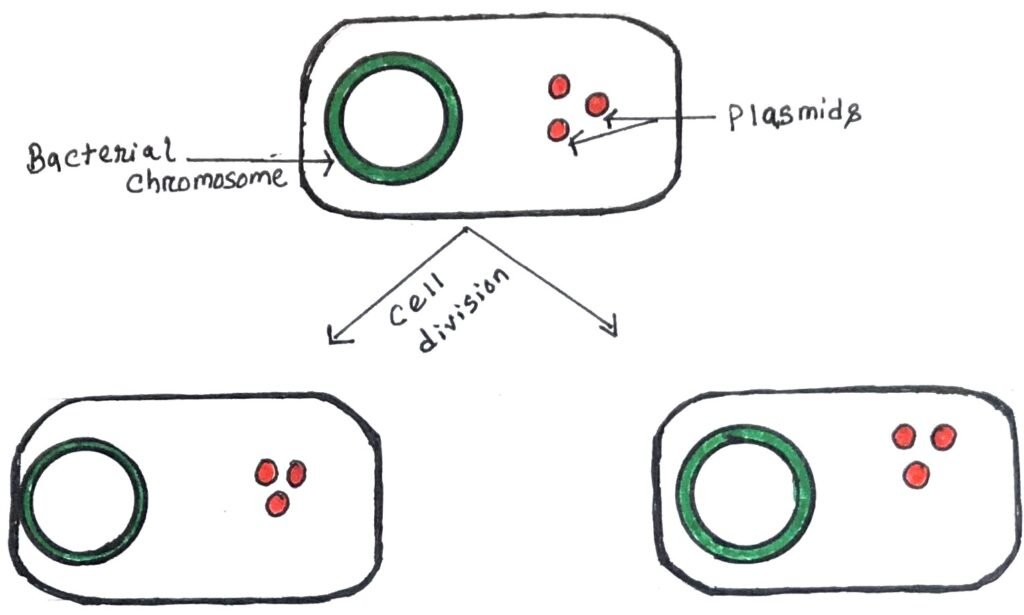
A plasmid is a circular DNA molecule that can replicate independently of the chromosomal DNA. Plasmids are considered replicons, capable of autonomous replication within a suitable host. Plasmids contain an origin of replication (ori), allowing them to multiply independently of the bacterial chromosome within the cell (Figure 1).
Some plasmids (integrative plasmids or episomes) can replicate by integrating into the bacterial chromosome (figure 2).
Unlike viruses, plasmids are “naked” DNA, meaning they do not encode the proteins required to package their genetic material for transfer to a new host. However, certain plasmids encode the sex pilus, which is necessary for their transfer. Plasmid transfer between hosts requires either direct mechanical transfer through conjugation or changes in host gene expression that enable the plasmid to be taken up through transformation. Smaller plasmids rely on the host cell’s DNA replication enzymes for their replication, while larger plasmids often carry genes that encode specific replication enzymes.

Two features of plasmids are important in cloning: size and copy number. A plasmid size of less than 10 kb is preferable for use as a cloning vector. The copy number indicates how many copies of a plasmid are typically present in a single bacterial cell, with each plasmid having a characteristic value that ranges from 1 to 50.
Types of Plasmids
Plasmids are categorized into two groups, i.e., conjugative plasmids and non-conjugative plasmids. Bacterial plasmids frequently contain antibiotic-resistance genes that help bacteria defend against antibiotics such as ampicillin. In genetic engineering, these genes act as selective markers, enabling scientists to verify whether the plasmid has been successfully inserted into a bacterial cell.
Conjugative plasmids can facilitate sexual conjugation between bacterial cells. This process is driven by a specific set of transfer (tra) genes, not found in non-conjugative plasmids. Tra genes encode proteins that facilitate the formation of the sex pilus, which is a structure that connects two bacterial cells, allowing the transfer of the plasmid from the donor to the recipient cell. This process enables the spread of genetic material, such as antibiotic-resistant genes, between bacteria.
However, under certain conditions, a non-conjugative plasmid can be co-transferred with a conjugative plasmid if both are present in the same cell. A single cell can harbor multiple plasmids; for instance, E. coli has been observed to contain up to seven types of plasmids simultaneously.
Plasmid Classification
Plasmids are classified into five main types:
- Fertility (F) Plasmids: These plasmids, like the F plasmid in E. coli, contain tra genes that enable them to facilitate conjugation between bacterial cells.
- Resistance (R) Plasmids: R plasmids carry genes that give resistance to antibiotics such as chloramphenicol and ampicillin, as well as toxic substances like mercury. They also help bacteria produce pili. These plasmids are particularly significant in clinical microbiology, as their spread can significantly impact the treatment of bacterial infections.
- Col Plasmids: Col plasmids have genes that encode bacteriocins, which are proteins capable of killing other bacteria. An example is the ColE1 plasmid found in E. coli. Bacteriocins are proteinaceous toxins, produced by bacteria to inhibit or kill closely related or competing bacterial strains. They are antimicrobial peptides that allow the producing bacteria to outcompete other microorganisms in the same environment. Bacteriocins are typically encoded by specific genes found on plasmids, such as Col plasmids in E. coli.
- Degradative Plasmids: These plasmids allow bacteria to break down and metabolize unusual substances, such as toluene and salicylic acid. An example is the TOL plasmid found in Pseudomonas putida.
- Virulence Plasmids: Virulence plasmids convert bacteria into pathogens. For instance, the Ti plasmids of Agrobacterium tumefaciens cause crown gall disease in dicotyledonous plants.
Bacteriophages
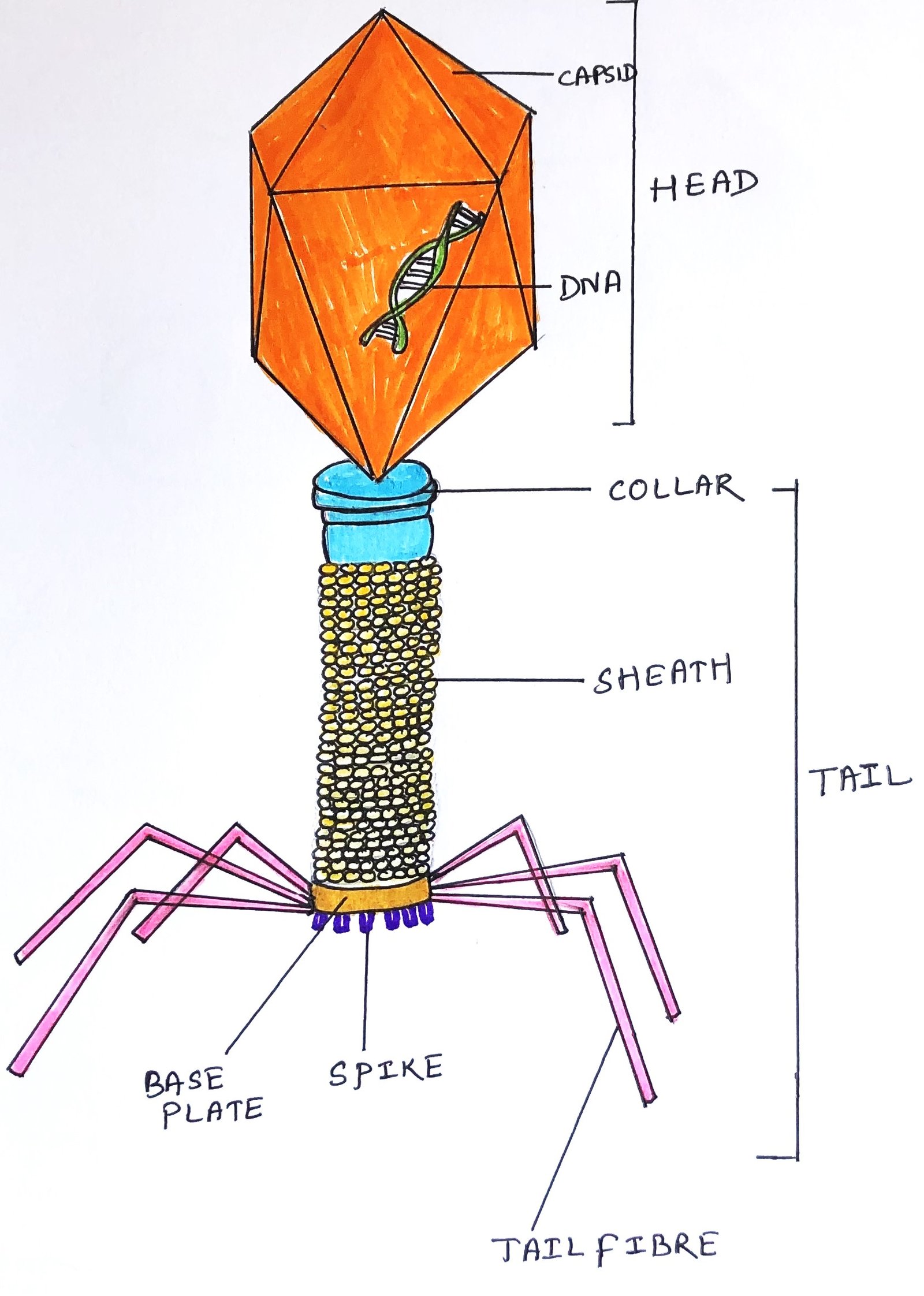
Viruses that specifically infect bacteria are known as bacteriophages. They all feature a head structure, which can vary in size and shape—some have an icosahedral shape while others are filamentous. The head is encased in a protective coat called a capsid, made of multiple copies of one or more proteins. The nucleic acid inside the head contains several genes for the phage’s replication. The head and tail are connected by a collar (figure 3). The tail is a hollow tube that delivers the nucleic acid during the infection. The tail varies in length among phages, and some lack a tail.

To initiate infection, the phage particle attaches to the outer bacterial wall and inserts its DNA chromosome into the cell. Genes on the phage chromosome encode specific phage enzymes, which help replicate the phage DNA molecule. Other phage genes direct the synthesis of the protein components of the capsid. Thus, new phage particles are assembled and released from the bacterium.
Lytic Cycle and Lysogenic Cycle
Bacteriophages can engage in either a lytic or lysogenic cycle, with some viruses capable of both. The T4 phage follows a lytic cycle where it undergoes rapid replication inside a bacterial cell, eventually causing it to burst open (lyse) and die. This lysis releases newly formed phages, which can then infect other bacterial hosts. Lytic phages are generally preferred for phage therapy. In some cases, lytic phages might undergo lysis inhibition, where the release of new phages is delayed if the concentration of phages outside the cell is high.
In contrast, the lysogenic cycle does not immediately destroy the host cell. Temperate phages can undergo lysogeny and integrate their viral genome into the host’s DNA. Here, it replicates along with the host’s genome without causing damage or forming a plasmid. The virus remains dormant until the host’s conditions worsen, such as during nutrient scarcity. At that point, the dormant phages, now known as prophages, activate and trigger the lytic cycle, leading to the host cell’s lysis. In the lysogenic cycle, the host cell remains alive and reproduces. Thus, the virus is passed on to the cell’s descendants.
Lambda (λ) Phage
The lambda phage is a virus composed of a head containing double-stranded linear DNA and a tail that may have tail fibers. This phage recognizes and attaches to its host, E. coli, triggering the injection of its DNA from the head through the tail into the bacterial cell’s cytoplasm. Typically, this initiates a “lytic cycle,” where the lambda DNA is replicated multiple times, and the genes responsible for the phage’s head, tail, and lysis proteins are expressed. This results in the assembly of many new phage particles inside the cell. It is followed by cell lysis, which releases the newly formed virions into the environment.
However, under certain conditions, the phage DNA can integrate into the host cell’s chromosome, entering the lysogenic pathway. The λ DNA molecule, which is 49kb in size, has been extensively studied through gene mapping and DNA sequencing techniques. A notable feature of the λ genetic map is that functionally related genes are clustered on the genome. This clustering is crucial for constructing λ-based cloning vectors, which is also significant for regulating the expression of the λ genome, as it allows for coordinated activation or repression of gene groups rather than individual genes.
Different Forms of Lambda (λ) DNA
The linear form of the λ DNA molecule consists of two complementary DNA strands. However, at each end of the molecule, there is a short 12-nucleotide sequence where the DNA is single-stranded. These single strands are complementary and can pair with each other to form a circular, fully double-stranded molecule.
These complementary single strands are often called ‘sticky’ or ‘cohesive’ ends because their base pairing can connect the two ends of a DNA molecule. The cohesive ends of λ DNA are known as cos sites, which enable the linear DNA injected into the cell to circularize. It is a necessary step before integration into the bacterial genome. The cos sites also play a crucial role after the prophage is excised from the host genome. At this stage, numerous λ DNA molecules are generated through the rolling circle replication mechanism, in which a continuous DNA strand is synthesized from the template molecule.
M13- Filamentous Phage
It is a filamentous bacteriophage containing a circular single-stranded DNA (ssDNA), which is made up of 6,407 nucleotides. The DNA is surrounded by approximately 2,700 copies of the major coat protein P8. It is capped at both ends by five copies of two distinct minor coat proteins: P9, P6, and P3. Although infection by filamentous phages like M13 is non-lethal, it leads to turbid plaques in E. coli and slows the growth rate of infected cells. M13 is a non-lytic virus, and its plasmids are commonly used in recombinant DNA techniques. The virus has also been heavily researched for its potential applications in nanostructures and nanotechnology.
Phage Life Cycle
The viral life cycle generally starts with infection, followed by viral genome replication, assembly of new viral particles, and release of these particles from the host cell. Filamentous phages such as M13 infect E. coli using the F pilus, with the phage’s p3 protein tip binding to the TolA protein on the bacterial pilus.
After attachment, the phage genome is delivered into the bacterial cytoplasm, where host proteins convert the single-stranded DNA genome into a double-stranded replicative form (“RF”). This replicative form serves as a template for phage gene expression and is not integrated into the bacterial genome. Instead, the phage DNA replicates within the host cell until over 100 copies are produced. Bacterial division produces daughter cells. Each daughter cell inherits copies of the phage genome. This genome continues to replicate, maintaining its presence within the cell.
The Factors Make M13 a Suitable Cloning Vector
A potential vector needs a small genome, which makes it befitting for cloning. The small genome size of M13 makes it a suitable cloning vector. The replicative form of M13 is double-stranded and can be used as a plasmid for experimental purposes. M13 can be easily prepared from a culture of E.coli cells and reintroduced by transfection. Single-stranded cloned genes obtained from M13-based vectors can be used in DNA sequencing and in vitro mutagenesis.
Conclusion
A cloning vehicle transports the gene into the host cell and replicates it. Plasmids and bacteriophages act as vectors for transporting genes. The vector carries the foreign DNA as the recombinant DNA. A plasmid is a circular DNA molecule that can replicate independently of the chromosomal DNA. Plasmids are considered replicons, capable of autonomous replication within a suitable host.
Plasmids can be categorized into two groups, i.e., conjugative and non-conjugative plasmids. They can be classified into five groups. They are fertility plasmids, resistance plasmids, virulence plasmids, col plasmids, and degradative plasmids.
Bacteriophages are the viruses that specifically infect bacteria. Some of them have an icosahedral shape while others are filamentous. Bacteriophages can engage in either a lytic or lysogenic cycle, with some viruses capable of both. The Lambda phage is a virus composed of a head containing double-stranded linear DNA and a tail that may have tail fibers. This phage recognizes and attaches to its host, E. coli, triggering the injection of its DNA from the head through the tail into the bacterial cell’s cytoplasm. M13 is a filamentous bacteriophage containing circular single-stranded DNA (ssDNA). It consists of 6,407 nucleotides. Its DNA is surrounded by approximately 2,700 copies of the major coat protein P8. The small genome size of M13 makes it a suitable cloning vector. The replicative form of M13 is double-stranded and can be used as a plasmid for experimental purposes.
You may also like:
- Cloning vectors based on E.coli plasmids
- Cloning vectors based on bacteriophages and cosmids
- Application of gene cloning in producing recombinant proteins

I, Swagatika Sahu (author of this website), have done my master’s in Biotechnology. I have around fourteen years of experience in writing and believe that writing is a great way to share knowledge. I hope the articles on the website will help users in enhancing their intellect in Biotechnology.