
In this article, I briefly describe the metabolism of fuel in fasting and starvation.
Hormonal and Metabolic Adaptations
Animals can withstand prolonged periods of starvation by adopting a comprehensive program of hormonal and metabolic adaptations. This type of adaptation mainly provides glucose for the brain and fat-derived fuels for most of the other tissues of the body. This is achieved through a well-orchestrated pattern of tissue-specific fuel metabolism. Hormones involved in metabolism are responsible for regulating fuel utilization. This includes the storage of any excess fuel and the appropriate mobilization of these stored fuels during the time of requirement. Metabolic hormones maintain a constant blood glucose level. This is done by the combined action of hormones insulin, glucagon, epinephrine, and cortisol on metabolic processes in many body tissues, especially in the liver, muscle, and adipose tissues.
The hormones epinephrine, norepinephrine, cortisol, and glucagon oppose the action of insulin and protect the body against hypoglycemia. When blood glucose concentration falls, the body defends it by first decreasing insulin secretion and then by releasing glucagon. When glucose homeostasis in the body fails, it leads to either hypoglycemia or hyperglycemia. Both conditions are dangerous and have damaging consequences.
The Metabolism of Glucose in Animals
The normal flow of nutrients in the body depends upon the appropriate amount of food. The immediate result of food deprivation is decreased intake of glucose. Glucose plays a vital role in the survival of animals. Certain cells, i.e., erythrocytes, cells of the renal medulla, and cells of the central nervous system, have an absolute requirement for glucose amounting to approximately 180 g/d. After the intake of a meal, the blood glucose level is decreased slightly in the first few hours. So, tissues receive glucose released from liver glycogen. However, after 24 hours of a meal, blood glucose has fallen further, causing a decrease in insulin secretion and an increase in glucagon secretion.
These hormonal signals mobilize fatty acids from triglyceride stores in adipose tissue. The stored fatty acids are used as an alternative fuel to glucose in muscles and the liver. This alternative fuel for energy production lowers the requirements for glucose, thus decreasing the demand for more glucose during the absence of input via feeding. Many adaptations in intracellular glucose metabolism occur, resulting in an inhibition of glucose-utilizing pathways and stimulation of glucose-producing pathways.
The process of glycogenolysis and gluconeogenesis helps to maintain the plasma glucose concentrations. In glycogenolysis, glycogen rapidly breaks down to glucose-1-phosphate and glucose. The amount of glucose released is insufficient to sustain the energy needs of the whole body for more than a short period. The liver and the kidneys synthesize glucose from different carbon sources via the process of gluconeogenesis. Glucose is synthesized primarily from glycerol, lactate, pyruvate, and certain amino acids, particularly alanine (Figure 1). Skeletal muscle releases lactate and pyruvate. Lactate arises from the reduction of tissue pyruvate.

The Cori Cycle
Carl Cori and Gerty Cori, the discoverers of the Cori cycle( Lactic acid cycle), refer to a metabolic pathway. In this pathway, lactate produced by anaerobic glycolysis in the muscles moves to the liver and is converted to glucose. The converted glucose then returns to the muscles and is converted back to lactate (Figure 2). When the rate of pyruvate production from glycolysis exceeds the glucose oxidation by the mitochondria, it results in lactate production. Therefore, for increased lactate production, mitochondrial glucose oxidation must decrease, which results in increased gluconeogenesis. The breakdown of triglycerides in adipose tissue produces glycerol. Proteins undergo breakdown in the liver and peripheral tissues to produce amino acids.

The Activity of the Pyruvate Dehydrogenase Complex (PDH) during Starvation
The pyruvate dehydrogenase complex (PDH complex) is the primary regulator of glucose oxidation in mammalian cells. At the enzymatic level, the pyruvate dehydrogenase (PDH) complex catalyzes the first irreversible reaction in the mitochondrial oxidation of glucose. The increased acetyl-CoA/CoA ratio and expression of pyruvate dehydrogenase kinase 4( PDK4) cause lipids to lower the PDH complex activity. The lipid-activated transcription factor, peroxisome proliferator-activated receptor-α (PPAR-α), plays a pivotal role in the cellular metabolic response to fatty acids.
During starvation and diabetes, the pyruvate dehydrogenase complex (PDH complex) is inactivated in many tissues to conserve three-carbon compounds for gluconeogenesis. The pyruvate is oxidized by the PDH complex in the presence of NAD+(oxidized form of nicotinamide adenine dinucleotide )and CoA( coenzyme A) to form acetyl-CoA, NADH, and CO2. The PDH complex regulates glucose oxidation, which is important to glucose homeostasis.
The activity of the PDH complex determines whether pyruvate is oxidized to CO2 and water or is converted to lactate via lactate dehydrogenase. To decrease the activity of the PDH complex, an increase in the extent of PDH complex phosphorylation is caused in part by increased pyruvate dehydrogenase kinase (PDK) activity. By reversible phosphorylation, the activity of the PDH complex can be decreased. Increased phosphorylation results in a decreased flux of glucose through the PDH complex. The activity of the PDH complex is tightly controlled by a phosphorylation/dephosphorylation cycle. The extent of phosphorylation of the complex is determined by the relative activities of the pyruvate dehydrogenase kinase (PDK) versus the pyruvate dehydrogenase phosphatase (PDP).
The Amino Acid Alanine- A Major Substrate for Gluconeogenesis
Amino acids represent an important precursor for gluconeogenesis. Most amino acids are released from muscle in proportion to their concentration in muscle proteins. However, the amino acids alanine and glutamine are released in greater amounts than their concentration in muscle proteins. This implies that de novo synthesis of alanine and glutamine occurs in skeletal muscle. The amino acid alanine is utilized by the liver and kidneys as a major substrate for gluconeogenesis. Felig and coworkers proposed a glucose-alanine cycle as a means of transferring amino nitrogen from muscle to liver and kidney. According to this hypothesis, glucose taken up by the muscle is metabolized to pyruvate. The pyruvate, instead of being oxidized via the PDH (pyruvate dehydrogenase) complex, serves as a nitrogen acceptor, with alanine being formed via transamination from glutamate. The alanine released from muscle recirculates to the liver, where it is converted back to glucose.
The Metabolism of Fuel in the Liver during Prolonged Fasting and Starvation
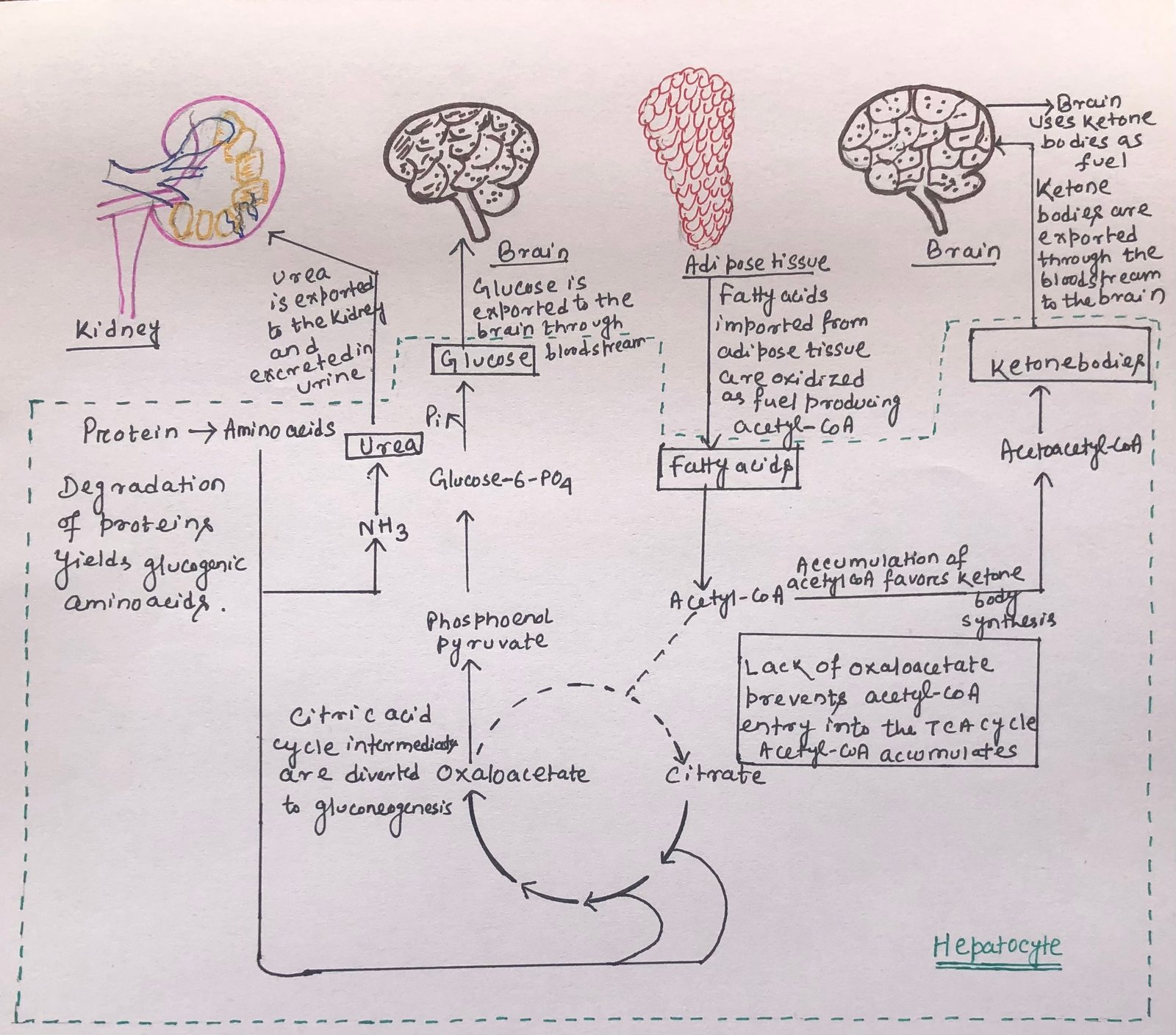
The liver degrades certain proteins to provide glucose for the brain. The non-essential amino acids are transmitted or deaminated, and the extra amino groups are converted to urea. The urea is exported via the bloodstream to the kidney and excreted (Figure 3). In the liver, the carbon skeleton of glucogenic amino acids gets converted into pyruvate or intermediates of the citric acid cycle.
These intermediates, along with the glycerol (derived from triacylglycerols) in adipose tissue, provide the starting material for gluconeogenesis in the liver. So, in this way, they produce glucose for the brain. However, the citric acid cycle intermediates, used for gluconeogenesis, depletes oxaloacetate, thus prohibiting the entry of acetyl-CoA into the citric acid cycle. The process of fatty acid oxidation produces acetyl-CoA, which favors the formation of acetoacetyl-CoA and ketone bodies in the liver (Figure 3). After a few days of fasting, the level of ketone bodies in the blood rises. It is because of the transportation of fuel from the liver to the heart, skeletal muscle, and brain. Thus, the use of these alternative fuels decreases the demand for glucose.

Acetyl-CoA allosterically inhibits pyruvate dehydrogenase and stimulates pyruvate carboxylase. Thus, acetyl-CoA stimulates the conversion of pyruvate to oxaloacetate by inhibiting its production from pyruvate.
In a normal adult, the triacylglycerols stored in the adipose tissue can render enough fuel to maintain a basal rate of metabolism for about three months. An obese adult has enough stored fuel to endure a fast of more than a year. The degradation of essential proteins starts after the utilization of the fat reserves, which leads to the loss of heart and liver function. Thus, this ultimately may lead to death.
The Action of the Hormone Epinephrine in Stress
During a stressful situation, neuronal signals from the brain trigger the release of the hormones epinephrine and norepinephrine from the adrenal medulla. The hormones epinephrine and nor-epinephrine, also called adrenaline, treat several conditions, e.g., cardiac arrest, anaphylaxis, and superficial bleeding. The hormones epinephrine and norepinephrine dilate the respiratory passages to facilitate the uptake of O2 and raise the blood pressure. Thus, they promote the flow of O2 and fuels to the tissues.
The hormone epinephrine mainly acts upon muscle, adipose tissue, and liver tissues. It stimulates the conversion of liver glycogen to blood glucose. Thus, generating the fuel for anaerobic muscular work by activating the enzyme glycogen phosphorylase and inactivating glycogen synthase. It also promotes the anaerobic breakdown of muscle glycogen by the fermentation of lactic acid, thus stimulating glycolytic ATP formation. The hormone epinephrine activates perilipin and triacylglycerol lipase, thus stimulating fat mobilization in adipose tissue.
The Action of the Hormone Cortisol in Stress
The hormone cortisol is the primary glucocorticoid in humans, and it maintains critical processes during times of stress. The hormone acts on muscle, liver, and adipose tissue to supply the organism with fuel to withstand stress. It alters metabolic, endocrine, nervous, cardiovascular, and immunologic systems to promote survival under stress.
Adrenocorticotropic hormone (ACTH), the principal hormone stimulating cortisol synthesis and secretion, is synthesized within the anterior pituitary. Pituitary ACTH release is stimulated primarily by corticotropin-releasing hormone. Corticotropin-releasing hormone is synthesized in neurons within the paraventricular nucleus of the hypothalamus.
During chronic stress, cortisol works to make fuel available. The hormone increases the concentration of fatty acids in the blood, and glycerol is released, which is available for gluconeogenesis. As a major catabolic effect, cortisol facilitates the conversion of protein in muscles and connective tissue into glucose and glycogen. For this, cortisol stimulates the breakdown of muscle protein to increase the plasma amino acid levels, which can then be used for gluconeogenesis in the liver.
In adipose tissue, cortisol increases the release of fatty acids from stored triacylglycerols. The fatty acids are exported to other tissues to serve as fuel. The glycerol is used for gluconeogenesis in the liver. In the liver, cortisol stimulates the synthesis of the key enzyme PEP carboxykinase, thus promoting gluconeogenesis. The glucose is stored in the liver as glycogen or exported immediately to tissues that need glucose for fuel. The hormone glucagon, like cortisol, stimulates the synthesis of the enzyme PEP carboxykinase, whereas insulin has the opposite effect. Thus, the effects of cortisol counterbalance the effects of the hormone insulin. These metabolic changes mainly restore blood glucose to its normal level and increase glycogen stores. In this way, they are ready to support the fight or flight response commonly associated with stress.
Conclusion
The metabolism of fuel in fasting and starvation includes an important role played by some hormones. Metabolic hormones maintain a constant blood glucose level. This is done by the combined action of hormones insulin, glucagon, epinephrine, and cortisol on metabolic processes in many body tissues, especially in the liver, muscle, and adipose tissues.
Glucose plays a vital role in the survival of animals. Certain cells, i.e., erythrocytes, cells of the renal medulla, and cells of the central nervous system, have an absolute requirement for glucose. When blood glucose concentration falls, the body defends it by first decreasing insulin secretion and then by releasing glucagon.
The liver degrades certain proteins to provide glucose for the brain. The non-essential amino acids are transmitted or deaminated, and the extra amino groups are converted to urea. The urea is exported via the bloodstream to the kidneys and excreted. In a normal adult, the triacylglycerols stored in the adipose tissue can render enough fuel to maintain a basal rate of metabolism for about three months. An obese adult has enough stored fuel to endure a fast of more than a year.
The hormone epinephrine mainly acts upon muscle, adipose tissue, and liver tissues. It stimulates the conversion of liver glycogen to blood glucose. The hormone cortisol is the primary glucocorticoid in humans, and it maintains critical processes during times of stress. The hormone acts on muscle, liver, and adipose tissue to supply the organism with fuel to withstand stress.
You may also like:
- Maintenance of blood glucose level by the hormones insulin and glucagon
- Triacylglycerols: Energy storage, insulation, and health impacts of partial hydrogenation

I, Swagatika Sahu (author of this website), have done my master’s in Biotechnology. I have around fourteen years of experience in writing and believe that writing is a great way to share knowledge. I hope the articles on the website will help users in enhancing their intellect in Biotechnology.