
In this article, I briefly describe the parallelism between the intestinal and respiratory immune systems.
The Respiratory Tract
The respiratory tract is responsible for breathing and exchanging oxygen and carbon dioxide between the body and the environment. It is divided into the upper respiratory tract and the lower respiratory tract. It is exposed directly to the external environment, like the gastrointestinal tract. Our respiratory tract is lined with mucous membranes and cilia (tiny hair-like structures) that help trap dust and microbes, keeping the lungs clean.
The Upper and Lower Respiratory Tract
The upper respiratory tract includes the nose, nasal cavity, sinuses, pharynx (throat), and larynx (voice box). These structures help humidify the air we breathe and trap particles and pathogens.
The lower respiratory tract consists of the trachea (windpipe), bronchi, bronchioles, and lungs. The trachea is separated from the oral cavity by a flap called the glottis. The trachea branches into bronchi, and eventually into bronchioles, which ultimately dead-end in clusters of tiny sacs called alveoli. These small sacs are the site of gas exchange. Oxygen from the air passes into the bloodstream, while carbon dioxide, a waste product from metabolism, is expelled from the blood and breathed out.
Similarities Between the Walls of the Respiratory Tract and intestinal tract
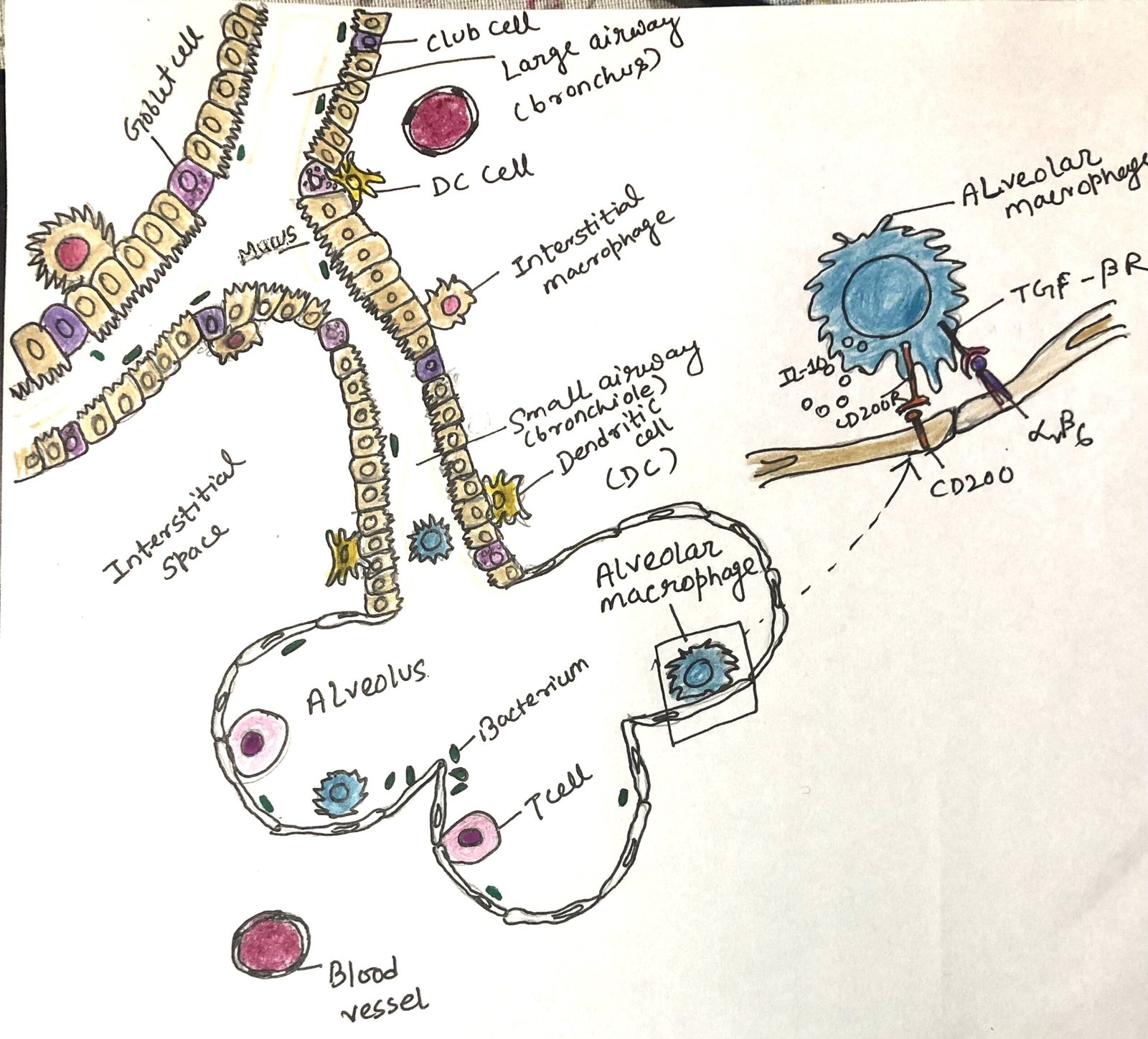
The epithelial layer of the respiratory tract and the intestinal tract share the same inhabitants as goblet cells, M cells, and the trans-epithelial process of antigen-presenting cells (Figure 1). The respiratory epithelium contains multipotent stem cells, like the intestinal epithelium. These multipotent stem cells replace damaged and dead epithelial cells.
The Unique Features of Respiratory Epithelial Cells
Cilia on the apical surfaces of most respiratory epithelial cells, along with mucus produced by goblet cells, create a mucociliary barrier. This barrier actively works to sweep away and remove microbes and particles that enter the airways. The respiratory epithelial layer consists of a unique secretory cell called the club cell, with many protective functions. The Club cells can also act as stem cells.
Specialized alveolar macrophages, also called dust cells, reside in the alveoli. The dust cells check out for infection in the lower airways and alveoli. They work along with respiratory epithelial cells and regulate the balance between tolerogenic and inflammatory responses.
The respiratory epithelial barrier maintains its functional integrity through different antimicrobial proteins and peptides secreted by respiratory mucosal cells and by secretory IgA. Antigen-specific IgA is produced in the same manner as in the intestine. The previous activation of innate and T lymphoid cells controls the production of the antibody.
The development of regulatory T cells takes place in the upper respiratory tract. Many commensal microbes residing here provide immune protection and spur tolerogenic activities. The same subsets of innate and adaptive immune cells found in the intestine are also found in the lamina propria of the upper respiratory tract. However, a healthy lower respiratory tract does not support a commensal microbial community.
NALT and BALT Support Immunity
Secondary lymphoid tissues and isolated follicles are found throughout the respiratory tract, with the nasal tissue having the most developed structures. The nasal-associated lymphoid tissue (NALT) is part of a lymphoid ring that includes the tonsils and adenoids. Structurally similar to Peyer’s patches in the intestines, NALT facilitates the activation of T and B cells, which are initiated by innate immune cells in the respiratory mucosa. In some animals, such as rabbits and cats, well-organized lymphoid tissue is located deeper in the lungs and is known as bronchus-associated lymphoid tissue (BALT). However, in humans and mice, this tissue is less organized and requires antigen exposure to fully develop, earning it the designation of inducible BALT.
Populations of Innate and Lymphoid Cells
The innate and lymphoid cells found in the intestine also reside in the lamina propria of the respiratory mucosa. These cells include CD103+ dendritic cells, CX3CR1+ macrophages, ILCs, regulatory T cells, IgA-secreting plasma cells, and more. Natural killer (NK) cells, also called cytotoxic ILC1 cells, are quite effective at locating and killing infected cells. These cytotoxic ILC1 cells are more abundant in the lung than in the intestinal lamina propria. ILC2s play an important role in maintaining the integrity of both the respiratory epithelium and the gut. The growth factor amphiregulin secreted by ILC2s interacts with epidermal growth factor (EGF) receptors on the epithelium, thus enhancing their health and growth.
Production of Regulatory T cells
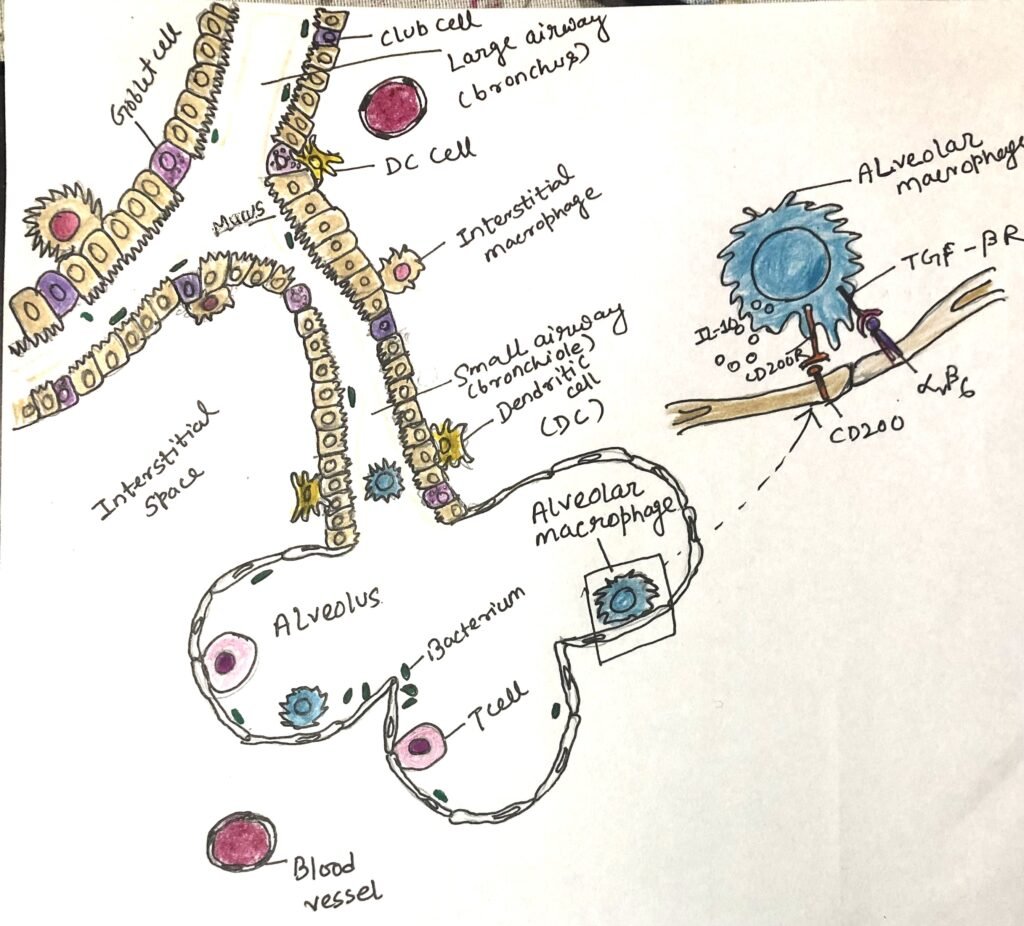
Upper airway epithelial and immune cells interact with commensal microbes to stimulate tolerogenic responses. The same kind of interaction takes place in the intestine. These interactions enhance the production and activity of regulatory T cells. B cells class switch to an anti-inflammatory IgA phenotype due to these interactions. Under normal, healthy conditions, alveolar macrophages in the lower airways receive anti-inflammatory signals from the epithelial cells, which express ligands for TGF-β (e.g., integrins αVβ6). These ligands interact with TGF- β receptors on the macrophages, promoting anti-inflammatory responses (Figure 1).
Induction of Type-1 and Type-2 responses
As in the intestine, a similar sequence of events happens in the respiratory immune system. Bacterial and viral microbes engage pattern-recognition receptors and antigen-presenting cells. This initiates a cascade of proinflammatory signals and cytokines. The cytokines IL-17, IL-23, and IL-1β program the antigen-presenting cells to induce type 1 response and activate ILC1, TH1, and TH17 cells and other subsets. Worms and allergens in the intestine stimulate epithelial cells to produce alarmins, including IL-25, IL-33, and TSLP. These trigger a type 2 response by activating ILC2 and TH2 cells to produce IL-4, IL-5, and IL-15.
Expression of Homing Receptors
Similar to the intestine, antigen-specific lymphocytes produced in lymph nodes that drain the lungs are prompted to express homing receptors, guiding them back to the respiratory tissues. One such receptor, CCR4, helps direct immune cells to the lungs, where its ligands, CCL17 and CCL22, are present.
Conclusion
The epithelial layer of the respiratory tract and the intestinal tract share the same inhabitants as goblet cells, M cells, and the trans-epithelial process of antigen-presenting cells. The respiratory epithelium contains multipotent stem cells, like the intestinal epithelium.
The nasal-associated lymphoid tissue (NALT) is part of a lymphoid ring that includes the tonsils and adenoids. Structurally similar to Peyer’s patches in the intestines, NALT facilitates the activation of T and B cells, which are initiated by innate immune cells in the respiratory mucosa.
The innate and lymphoid cells found in the intestine also reside in the lamina propria of the respiratory mucosa. These cells include CD103+ dendritic cells, CX3CR1+ macrophages, ILCs, regulatory T cells, IgA-secreting plasma cells, and more.
Upper airway epithelial and immune cells interact with commensal microbes to stimulate tolerogenic responses. The same kind of interaction takes place in the intestine. These interactions enhance the production and activity of regulatory T cells.
As in the intestine, a similar sequence of events happens in the respiratory immune system. Bacterial and viral microbes engage pattern-recognition receptors and antigen-presenting cells. This initiates a cascade of proinflammatory signals and cytokines. The cytokines IL-17, IL-23, and IL-1β program the antigen-presenting cells to induce type 1 response and activate ILC1, TH1, and TH17 cells and other subsets.
Similar to the intestine, antigen-specific lymphocytes produced in lymph nodes that drain the lungs are prompted to express homing receptors, guiding them back to the respiratory tissues.
You may also like:
- Intestinal Immunity Can Initiate Both Type 1 and Type 2 Immune Responses
- Various innate and adaptive cell types promote immune homeostasis in the intestine
- The Skin Immune System

I, Swagatika Sahu (author of this website), have done my master’s in Biotechnology. I have around fourteen years of experience in writing and believe that writing is a great way to share knowledge. I hope the articles on the website will help users in enhancing their intellect in Biotechnology.